Immunogenicity and Protection by DnaK and SpaA Recombinant Proteins Against Erysipelothrix Rhusiopathiae in a Murine Model
Keywords
Abstract
Background/Aims:
Swine erysipelas is a disease caused by Erysipelothrix rhusiopathiae , a Gram-positive bacillus, which has great economic importance because it leads to the loss of the swine herd. To control this disease, animals are immunized with a cellular vaccine of killed or attenuated E. rhusiopathiae , but even with herd vaccination, cases of swine erysipelas outbreaks have been reported in the United States, China and Japan, leading to the search for other antigenic components of the bacteria that may promote greater protection against E. rhusiopathiae . The surface protein SpaA from E. rhusiopathiae has been shown to be a candidate to constitute a subunit vaccine, since it has already been reported to induce a host immune response against the bacterium. DnaK, a hsp70 molecular chaperone, also seems to be a good candidate in the composition of a vaccine, as it has been demonstrated to be an antigenic protein of the bacteria.Methods:
This work evaluated the immunogenicity and protection induced by the E. rhusiopathiae SpaA and DnaK recombinant proteins in a murine model, by intramuscular administration to mice with two doses of 100 µg at 21-day interval between them. The candidate proteins were tested either separately and together, compared with the commercial vaccine and the non-vaccination condition, and mice were challenged with a virulent strain of E. rhusiopathiae. Serum was collected to assess the produced antibodies and peripheral blood cells, whereas spleen and kidney tissues were assayed for E. rhusiopathiae presence by colony counting.Results:
A survival curve of the animals was performed, which confirmed the protection induced by the proteins. IgG antibodies increased in the animal serum inoculated with the proteins when compared to the control, and a significant delay in disease symptoms was observed.Conclusion:
These results suggest that E. rhusiopathiae DnaK and SpaA are immunogenic in mice and interfere with the disease development.Introduction
In the last 40 years, global pig meat consumption has grown at a rate of 1.52% per year, with China being the largest producer of pork, Japan the largest importer and the United States the largest world exporters. Brazil is the 4th largest exporter of pig meat, accounting for 8% of world trade [1, 2].
Swine erysipelas, when uncontrolled, is an economically significant disease, capable of affecting all stages of swine production through sudden death, septicemia and the development of arthritis and/or endocarditis in the chronic phase of the disease [3, 4]. The Gram-positive bacterium E. rhusiopathiae is the infectious agent of swine erysipelas, having several pathogenicity factors, such as the presence of a polysaccharide capsule on its cell surface that promotes resistance to the phagocytosis process, as well as enzymes and proteins that may be involved in the process of bacteria adherence to host cells [5-7]. From an economic point of view, erysipelas arthritis is one of the most important clinical forms of the disease, as it consists in a proliferative and non-suppurative form, which can progress in alive swine even in the absence of the bacteria. This condition causes harm to the animal development, in addition to the losses due to the devaluation of meat after slaughter [3, 8].
The mainstay of treatment for infections caused by E. rhusiopathiae is antibiotic therapy, such as penicillin and cephalosporin, but the constant use of an antibiotic can induce bacterial resistance making some strains less sensitive to this drug, as has already occurred with erythromycin, making treatment difficult [4, 9]. Thus, to prevent the emergence of resistant strains and avoid a large loss of swine and the devaluation of meat after slaughter, the prevention of the erysipelas through the use of vaccines has been shown to be the best alternative to control this disease [9, 10].
The currently used vaccines, composed of dead or attenuated E. rhusiopathiae cells, are able to control the disease, however, in recent years some countries have already reported outbreaks of swine erysipelas and also the development of the disease in swine, even after immunization [11, 12]. In the United States, in the summer of 2001, cases of increased erysipelas were reported in both vaccinated and non-vaccinated pigs, and in 2010 there were outbreaks of the disease reported in 6 swine operations [13, 14].
An outbreak of acute swine erysipelas was reported in Eastern China, in which seven other E. rhusiopathiae strains were isolated since the summer of 2013 [15]. In Japan, approximately 2000 cases of erysipelas in pigs are reported per year, with sporadic outbreaks of the disease reported from 2008 to 2019 mainly caused by the Met-203 type strain, suggesting that this strain might be widespread in the country [16-18]. Due to the high frequency of occurrence of these outbreaks, there is a suspicion that the vaccine composed of the attenuated bacterium may have its virulence reversed. However, it is difficult to prove that the strain present in the vaccine was the same responsible for the outbreak of disease, therefore vaccines composed by antigenic parts of the pathogenic agent could be a promising alternative to provide higher protection and safety [11, 19, 20].
Subunit vaccines are composed by purified fractions of the pathogen, such as proteins capable of inducing the immune response in the host even in the absence of the infectious agent [21]. An example is the surface protein SpaA from E. rhusiopathiae , which is considered an antigen with good potential for the design of protective vaccine for swine erysipelas [22, 23]. Furthermore, the hsp70 molecular chaperone known as DnaK can also be considered as a potential candidate for a subunit vaccine as it has been reported to be able to activate host immunity, and, as a heat shock protein, to act as a vaccine adjuvant although its protective activity has not been tested [24, 25]. Also, a previous immunoproteomic analysis performed by our research group (LBBMA- Department of Genetics and Evolution, UFSCar), with the extracellular fraction of E. rhusiopathiae could detect anti-DnaK antibodies in the sera of swine immunized with a commercial E. rhusiopathiae cellular vaccine (V. Marquioni, unpublished results).
This work aimed to analyze the immunogenicity and protective properties of the E. rhusiopathiae DnaK antigenic protein, and compare its role separately and together with the SpaA protein, already reported in literature as involved in the bacterium virulence and antigenicity.
Materials and Methods
Construction of pET28a_DnaK and pET28a_SpaA clones in ClearColi™ Bl21 DE3
E. rhusiopathiae NCTC11002 was used for genomic DNA extraction with the kit AxyPrepTM (Axygen), and DnaK was PCR amplified with the primers DnaK_For_NdeI (GGCGGCATATGAGTAAAGTTATTGGTATTG ) and DnaK_Rev_BamHI (CGGCGGATCCTAGTTTTTTTCTTCAAATTC ), which were designed based on genome sequence at GenBank® NCBI (National Center for Biotechnology Information, AP012027), whose underlined sequences are sites for the respective restriction enzymes. The DnaK is located at the ERH_RS02935 locus, between nucleotides 615078 and 616880 and the amplified fragment was ligated between the NdeI and BamHI sites of the pET28a(+) plasmid (Novagen) for expression in fusion with a 6-histidine tag (His-tag ). The construction is illustrated in Supplementary Material Fig. S1. For SpaA expression was used pET28a(+) vector containing the 1026-bp fragment coding for the SpaA antigenic region (268 to 1293 nt position), which was obtained previously by da Silva et al [26]..
Recombinants DnaK and SpaA were properly expressed in ClearColi™ (Lucigen) according to Supplementary Material and Methods 1.
Purification of rDnaK and rSpaA proteins
The proteins rSpaA and rDnaK were purified from the soluble and insoluble fractions respectively, (fractions were obtained by sonication, as described in Supplementary Material and Methods 2), by immobilized metal ion affinity chromatography (IMAC) [27], using for rDnaK a Poly-Prep® (Bio-Rad) column with 1 mL of Ni SepharoseTM 6 Fast Flow resin (GE Healthcare). After soluble fraction was added, column was washed 3 times with 5 times the column volume (CV) of this same buffer, and proteins were recovered by adding 1 CV of the same buffer containing imidazole of 2 mM to 2000 mM. rDnaK purified from the elution solution (5 mL) was concentrated to 1 mL by ultrafiltration with Amicon® Centrifugal Filter Unit (Merk Millipore) and molecular exclusion chromatography was performed using a Superdex 200 column (GE Healthcare) with a flow rate of 0.5 mL/minute. The recombinant protein was eluted in 3.5 mL Tris-NaCl buffer solution (50 mM Tris-HCl, 150 mM NaCl, pH 8) and concentrated to 1 mL.
rSpaA were solubilized from inclusion bodies under gentle agitation for 45 minutes at room temperature in 100 mM Tris-HCl, pH 8; 1% TritonX-100 and 3 M urea, being the solution centrifuged at 18, 000 x g for 30 minutes at 4 °C, and pellet washed twice in the same buffer. Pellet was agitated in a solubilization buffer (50 mM Tris-HCl, pH 8; 10 mM DTT; 8 M urea) for 20 hours at 10 °C. Soluble rSpaA was added to a 5 mL of nickel IMAC resin equilibrated with binding buffer (20 mM Na2HPO4.12H2O, pH 7.4; 500 mM NaCl; 20 mM imidazole; 6 M urea), and column was washed twice with 5 CV of this buffer with no imidazole (elution buffer). rSpaA was recovered in 5 mL of elution buffer with decreasing pH values of 7, 6.3 and 4.5, followed by concentration to 1 mL by ultrafiltration. All samples were analyzed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), using coomassie Blue-silver gel staining [28]. Quantification was performed using Protein Assay Dye Reagent Concentrate (Bio-Rad) [29].
Vaccinated and non-vaccinated swine sera
Three adult male pigs had their serum collected before vaccine application (day 0), and soon after, the same pigs were immunized with the Farrowsure® B Gold (Pfizer) vaccine, composed of live attenuated E. rhusiopathiae . After 21 days from immunization, serum from the same three pigs were collected (day 21), and they were again immunized with a second dose of the vaccine. The last collection of serum from these pigs was performed 15 days after the second immunization (day 36). Vaccination and serum collection were performed by Cripion Biotechnology (Andradina, Brazil), and serum was used for Western Blot and ELISA assays.
rDnaK and rSpaA Western Blot against swine sera
rDnaK and rSpaA were assayed using sera from immunized and non-immunized swine. Five µg of protein were submitted to SDS-PAGE and transferred to 0.45 µm nitrocellulose membrane in triplicate, using a Mini-PROTEAN® system (Bio-Rad) at 400 mA for 2 hours. After overnight blocking with 9% powdered milk, membranes were incubated for 2 hours with a 1:5000 dilution of the swine sera before immunization and after 15 days from the second immunization, followed by incubation for 1.5 hours with the 1:5000 dilution of the secondary Anti-swine IgG Antibody Conjugated to Peroxidase (KPL). Chemiluminescence was detected with the ClarityTM Western ECL Substrate kit (Bio-Rad) and documented using ChemiDoc™ system (Bio-Rad).
Mice immunization with the rSpaA and rDnaK and E. rhusiopathiae challenge
Sixty pathogen free (SPF) four-weeks old BALB/c female mice were obtained from the Central Biotery of University of São Paulo (Ribeirão Preto). Protocols were approved by the Ethics Committee for Animals Use (CEUA) at UFSCar (nº1247201120). As a reference, intramuscular application of 40 µL of the SUIVAC® ERY-IN (Dyntec) commercial vaccine of inactivated E. rhusiopathiae cells was used, a dose 50 times lesser than the commercially used in pigs (2 mL).
Inoculum for mice immunization was prepared with each recombinant protein (100 µg) or both (50 µg of each one) mixed with 40 µg of Al(OH)3 adjuvant (Dinâmica) and diluted in 100 µL of 1X PBS (137 mM NaCl, 10 mM Na₂HPO₄, 1.8 mM KH₂PO₄, 2.7 mM KCl, pH 7.4). Sixty mice were divided into 6 groups (G1 to G6, N=10/mice per group) (Fig. 1), being five from each group separated for the survival curve after E. rhusiopathiae challenge, performed with 100 µL of subcutaneous inoculation of 4000 cells of NCTC11002 strain. Blood, serum, spleens, and right kidney were collected from the remaining 30 animals after 3 days of challenge. Blood was collected from the mice brachial vein using a Pasteur pipette soaked in anticoagulant, and serum was obtained by centrifugation (7000 x g for 15 minutes). Animals were anesthetized by intraperitoneal anesthetic overdose containing 270 mg/kg of ketamine, Dopalen® (Ceva) and 30 mg/kg xylazine, Anasedan® (Ceva).
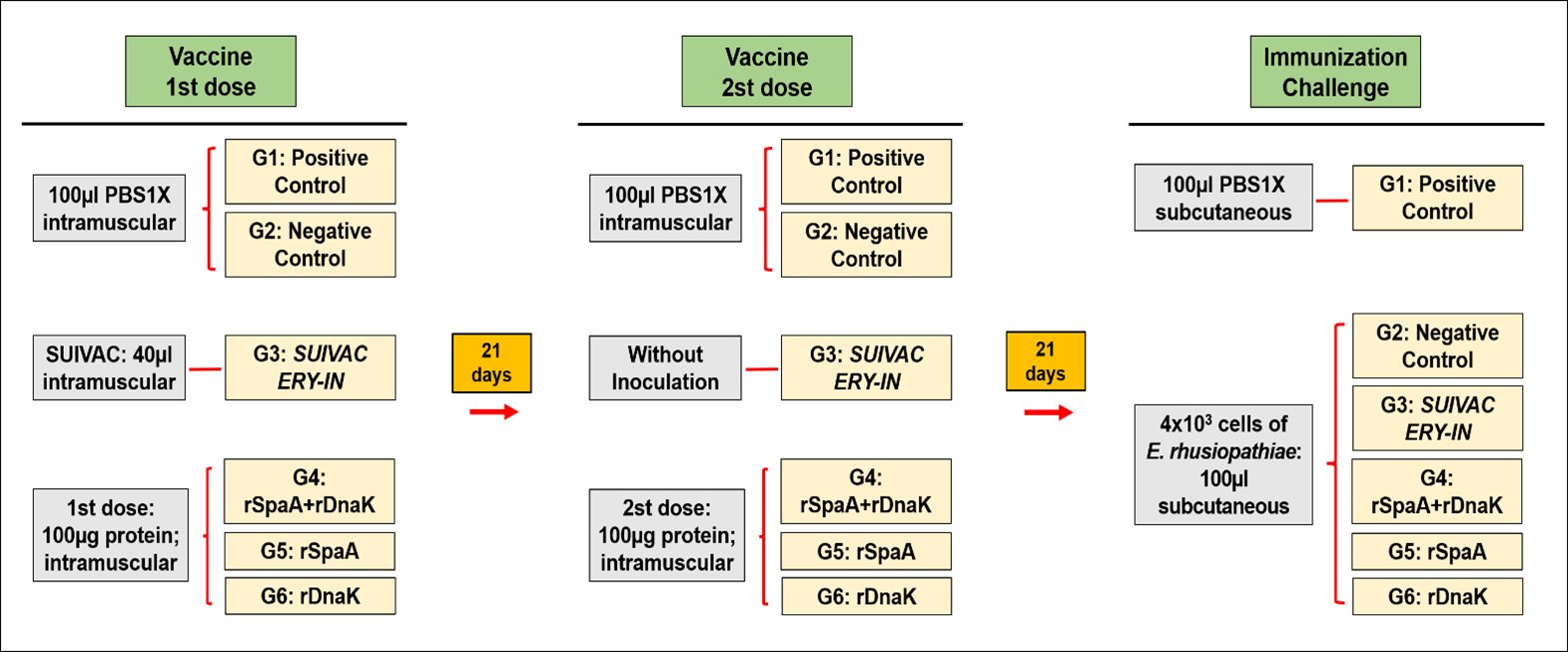
Fig. 1: Experimental design of the immunization trial. Six different groups were established. The mice were immunized intramuscularly with the purified recombinant proteins and with the comercial vaccine. The positive and negative control group were only injected with PBS 1X. Twenty-one days post injection, a second dose with the recombinant proteins were inoculated. After twenty-one more days, the challenge with the bacterium E. rhusiopathiae was carried out subcutaneous and the positive control were injected again with only PBS 1X. Three days after the challenge, the mice's blood was collected for further analyses.
IgG analysis in swine and mice serum by ELISA
Animal sera were analyzed by ELISA (Enzyme-Linked Immunosorbent Assay) for specific antibodies against rSpaA and rDnaK E. rhusiopathiae . The 96-well plates (Costar) were sensitized with 50 ng protein/well of purified rDnaK and/or rSpaA in carbonate buffer (87 mM NaHCO3; 15 mM Na2CO3 pH 9.5). After overnight incubation at 4 °C, each well was washed 3 times with 1X Phosphate-buffered saline (PBS) with Tween®20, blocked with 200 μL of 1% bovine serum albumin for 2 hours at 37 °C and washed again. For IgG analysis, 100 μL of serum diluted 1:10; 1:100; 1:1000 and 1:10000 were added followed by incubation at 37 °C for 1 to 2 hours. After washing, 100 µL of the 1:5000 dilution of the secondary anti-mouse IgG antibody (IgG Total; IgG1 and IgG2a) conjugated to Goat Anti-Mouse IgG Peroxidase Antibody (GenScript) were added. After incubation for 1 hour at 37 °C and washing, detection was performed by addition of 50 µL of 3, 3′,5, 5′‑tetramethylbenzidine (TMB) and incubation for 30 minutes in the dark at room temperature. 50 µL of 1 M H2SO4 were added, and the absorbance was read at 450 nm in a microplate reader (Bio-rad).
Mice peripheral blood count
Global count was performed in optical microscope using Neubauer chamber immediately after blood collection, and the differential cell count was performed using blood smears stained with the Rapid Panoptic Kit (Laborclin) prepared at the time of puncture. Differential count is obtained between the global count of each group and the count of 100 cells of the blood smear differentiated in mononucleated, neutrophils, and eosinophils [30].
E. rhusiopathiae biodistribution in the spleen and kidney tissues
The spleens and right kidneys of 3 animals from each group were removed and macerated separately in microtubes containing 1 mL of sterile PBS. Aliquots (100 µL) of 1:10 or 1:100 dilutions were plated on LB agar and incubated at 37°C. After 48 hours, Colony Forming Units (CFUs) were counted.
Survival Curve
Five mice from each of the 6 groups (G1 to G6) were monitored for 14 days to analyze the percentage of animals that survived or died after bacterial challenge, to build a survival curve [31]. All remaining animals were euthanized by an overdose of anesthetics containing 270 mg/kg of ketamine, Dopalen® and 30 mg/kg xylazine, Anasedan®.
Statistical Analysis
Quantitative data were analyzed by one-way ANOVA to compare means using the Prism® software (version 5.0, GraphPad Software). Individual differences between groups were identified using Dunn’s test. The results were expressed as mean ± standard error of the mean (SEM) and, in all analyses, 5% was adopted as the statistical significance limit (p < 0.05).
Results
Expression and Purification of rDnaK and rSpaA Proteins
The confirmation of pET28a+DnaK clone in a ClearColi colony was performed by digestion of the plasmid with restriction enzymes Nde I and Bam HI, which provided a 1.8 kb digested fragment, whereas the transformation of the pET28a+SpaA construct into ClearColi was validated by PCR colony, resulting in a 1.026 kb fragment amplification (Supplementary Material Fig. S2).
The rDnaK and rSpaA were properly expressed in endotoxin-free ClearColi, and purified from the soluble and insoluble fractions, respectively (Supplementary Material Fig. S3). The DnaK total sequence of 65 kDa [23] was expressed fused with 1 kDa His-Tag, which is coded by the pET28a vector. The rDnaK derived from this expression was mainly eluted by 250 mM imidazole during the purification by IMAC (Fig. 2), and to further purify the protein from imidazole and other contaminant proteins, molecular exclusion chromatography was performed, being rDnaK eluted between 60-80 nm between 43 and 75 mL of elution solution (Suplemmentary Material Fig. S4). The purified proteins were confirmed as the derivatives of the ClearColi clones by reacting the His-Tag present in the construct (Supplementary Material Fig. S5).
The native E. rhusiopathiae SpaA protein has a size of approximately 64 kDa [32], but In this work the SpaA was expressed as a partial sequence of 42 kDa [26, 33], fused to a 1 kDa His-Tag . The rSpaA was effectively solubilized before purification in the nickel column by IMAC, in which the recombinant protein passed through the column even before the decrease in the pH of the elution buffer, but fractions eluted at pH 4.5 showed a high purity of a protein with the expected size (42 kDa) for the expressed rSpaA (Fig. 3).
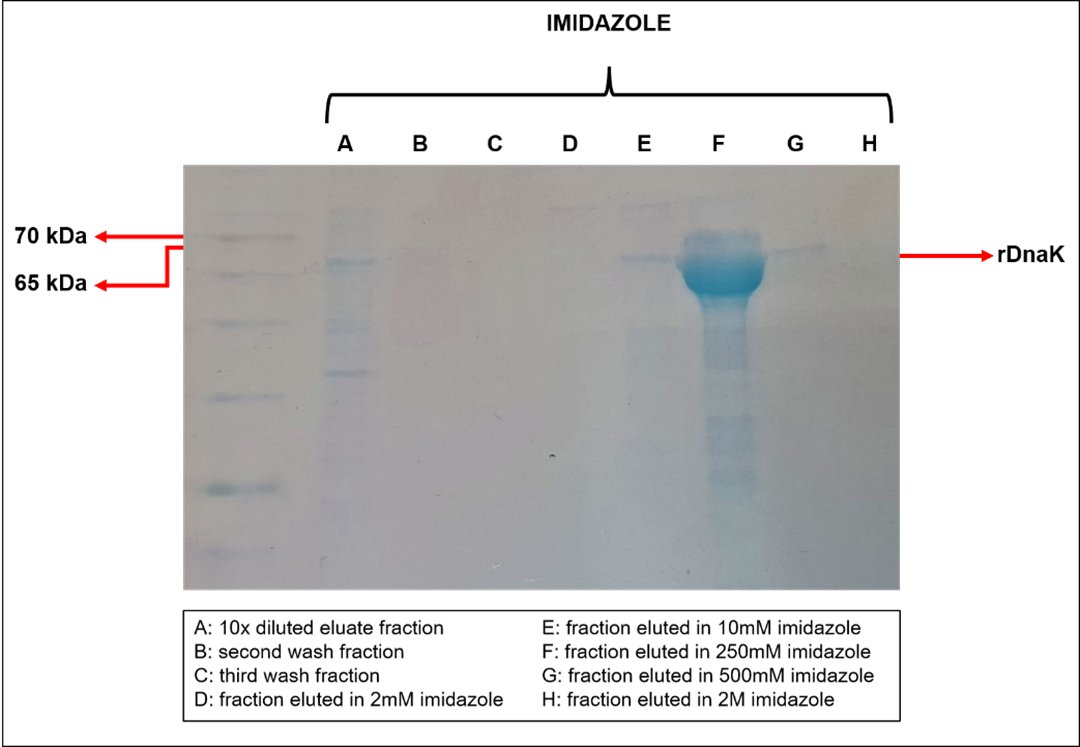
Fig. 2: SDS-PAGE analysis of rDnaK protein purification by IMAC with imidazole gradient elution. The rDnaK protein was induced at 20 °C and with 0.1 mM of IPTG, in 250 mL of LB medium, by the ClearColi expression system. Purification was performed with 1 mL of nickel resin and eluted with an imidazole gradient (2 mM; 10 mM; 250 mM; 500 mM and 2 M).
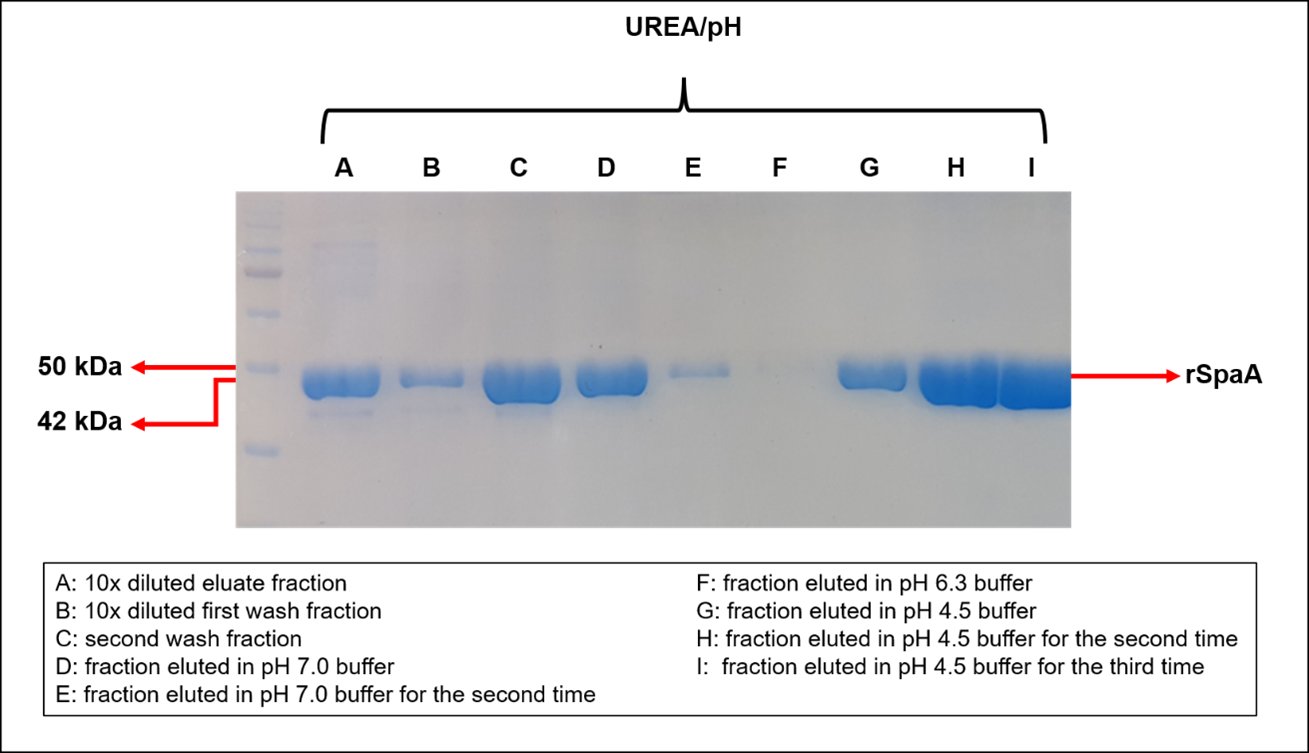
Fig. 3: SDS-PAGE analysis of rSpaA protein purification by IMAC with pH decrease elution. The rSpaA protein was expressed through the ClearColi expression system at 20 °C and with 0.1 mM of IPTG in 250 mL of TB medium, and after cell lysis it was solubilized with 8 M urea from the inclusion bodies. The rSpaA was purified on a column with 5 mL of nickel, and eluted by pH difference (7; 6.3 and 4.5) of the elution buffer.
Immunoreactivity of rSpaA and rDnaK proteins with swine serum
The purified proteins 42 kDa rSpaA and 65 kDa rDnaK reacted to antiserum from previously immunized pigs. Significant differences between the sera collected after first and second immunization were observed (Fig. 4). Post-immunized sera (especially in the 1:10 and 1:100 dilutions) presented increased anti-DnaK IgG antibodies in relation to the pre-immunized sera, mainly after the second dose of vaccine. In Fig. 7, swine sera (15 days after the second immunization) were used as primary antibodies to assess their reactivity against purified rSpaA and rDnaK.
The rSpaA protein showed low reactivity against swine sera collected before immunization, while rDnaK showed no detectable reactivity. However, the two proteins showed immunoreactivity with all sera collected after the second vaccination dose. Post-immunization sera of each swine presented different immunoreactivity to proteins, in which the serum from swines B and P showed approximately the same intensity of reaction against both rDnaK and rSpaA, being however swine P more reactive, whereas swine C was more reactive to rSpaA than rDnaK (Fig. 5).

Fig. 4: BoxPlot of Total IgG absorbance data against swine sera collected at different periods in different groups. P0 corresponds to the serum of pigs collected before immunization; P21 refers to the serum of pigs immunized 21 days after the first dose of the commercial vaccine; P36 indicates the serum collected 36 days after the second immunization with the commercial vaccine. *p<0.05 represents the significant difference between the groups.
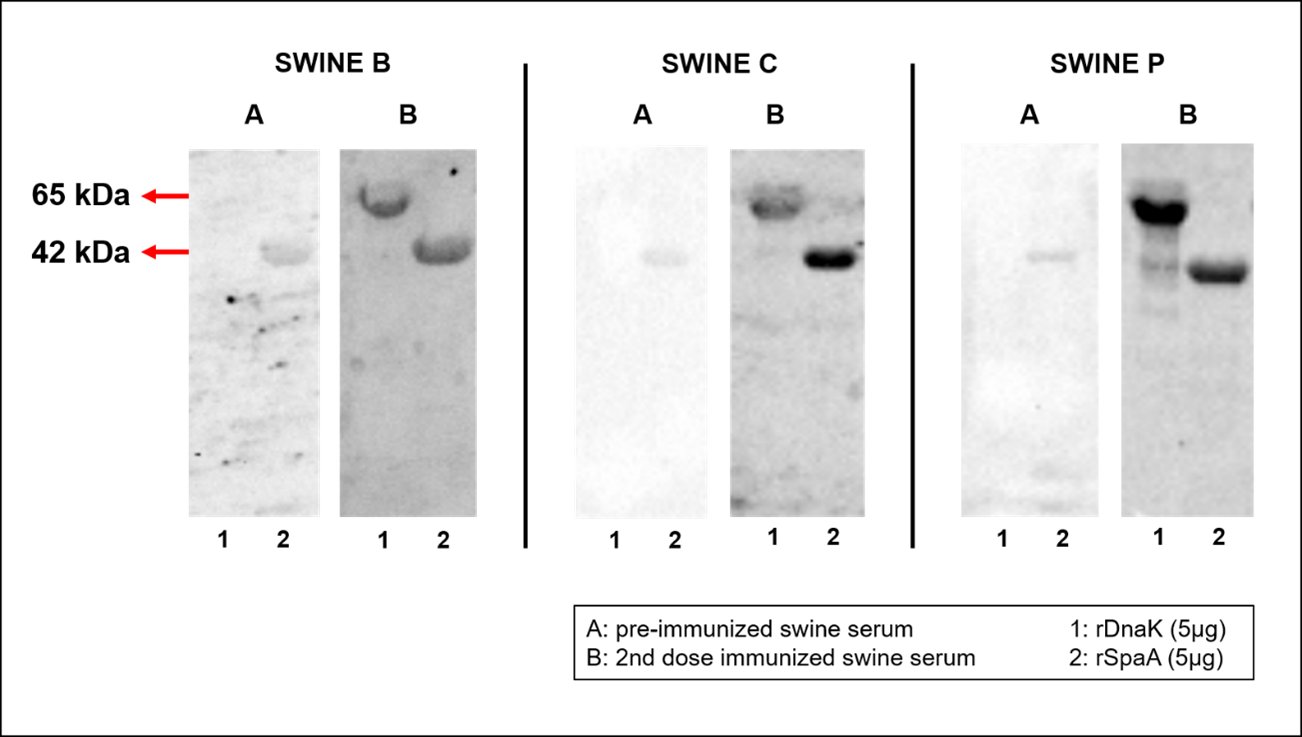
Fig. 5: Western blot of purified rSpaA and rDnaK recombinant proteins against sera from swine pre and post-immunized with the commercial vaccine against E. rhusiopathiae. Columns A represent the sera of the swine before being immunized with the commercial vaccine; and column B represent swine sera after the second dose of vaccine. The purified rDnaK and rSpaA proteins were equalized (5 µg) and represented by the numbers 1 and 2, respectively.
rSpaA and rDnaK immunogenic effect against E. rhusiopathiae
After 3 days of the challenge with the bacteria, some animals began to show symptoms of the disease, and before these animals died the serum of mice from all groups were collected. Both rSpaA and rDnaK showed to have the ability to induce an immune response in mice against the bacterium. The presence of anti-rSpaA and anti-rDnaK IgG total antibodies in the serum of the animals of the immunized groups in a higher amount than in the non-immunized two groups G1 and G2, indicates that the heterologous proteins rSpaA and rDnaK in G4, G5 and G6 were immunogenic, generating a specific humoral response in these 3 test groups (Fig. 6A).
There was no significant difference in the amount of IgG total or IgG1 produced among the groups G4, G5 and G6 (Fig. 6A and 6B). As for IgG2a antibodies, all immunized groups showed a significantly higher production of this antibody when compared to the control groups, but the group immunized with rSpaA showed a higher IgG2a than each one of the other vaccinated groups or even the group immunized with the commercial vaccine SUIVAC (Fig. 6C).
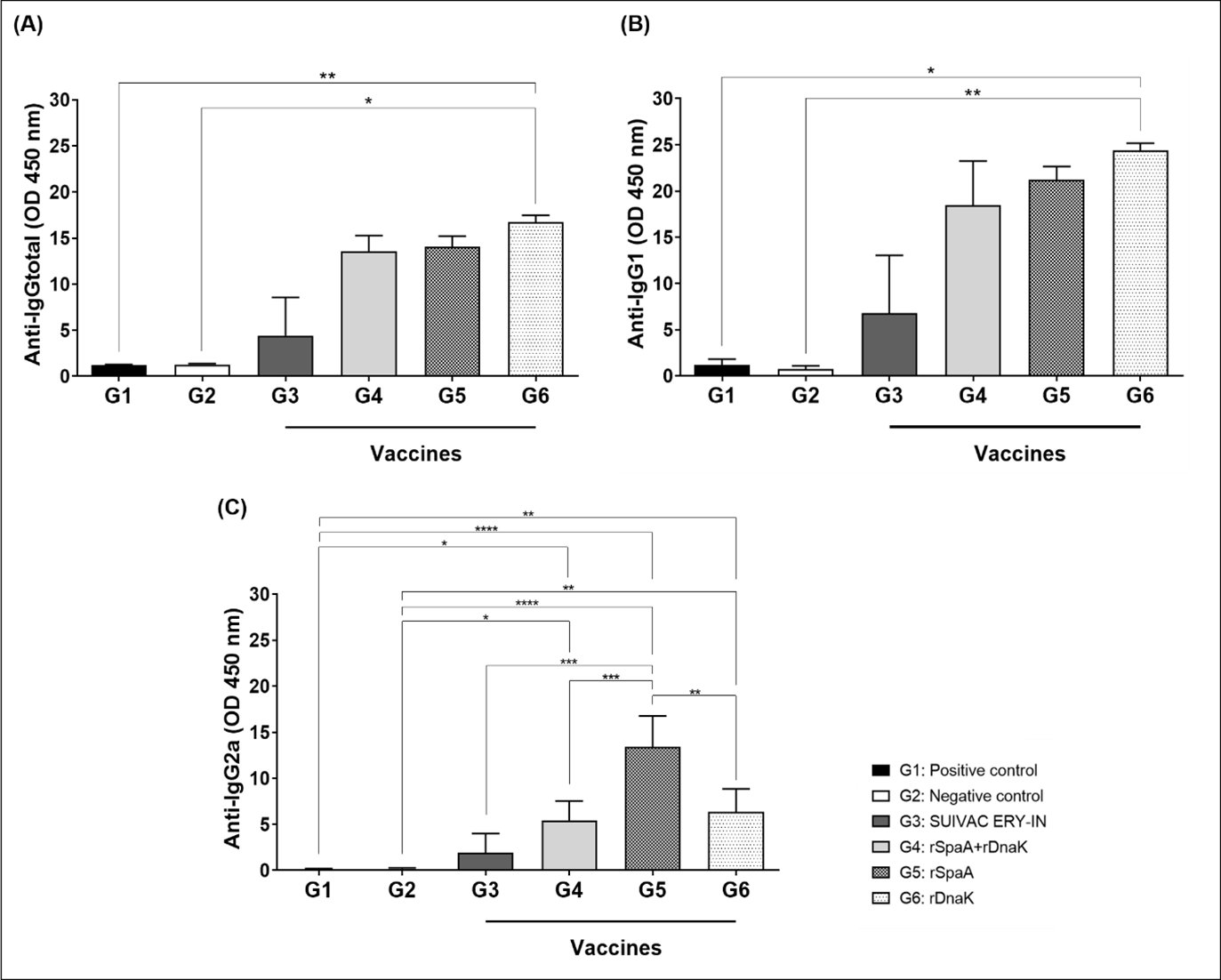
Fig. 6: Total IgG titers; IgG1 and IgG2a in the blood of isogenic mice, 3 days after infection. Subclasses of IgG antibodies in the serum of each 4 mice for group were indirectly quantified against the antigenic proteins rSpaA and rDnaK. (A) Indirect quantification of Total IgG. (B) Indirect quantification of IgG1. (C) Indirect quantification of IgG2a. Data were normalized, and represent the mean of 4 animals. *p<0.05 represents the significant difference between the groups.
Peripheral blood cell profile analysis after immunization
The blood of 4 mice from each group was analyzed, in which 100 cells, subdivided into mononuclear cells, neutrophils and eosinophils, were counted. Based on the data obtained, an analysis was performed to observe the difference in the profile of blood cells present in each group. There was no significant difference in the number of mononuclear cells between the vaccinated groups neither with the positive control. We found a difference in the number of cells between the immunized groups and the negative control, but only the group immunized with rSpaA showed a significant difference (Fig. 7A). In the neutrophil count, a greater number of cells was observed in the vaccinated groups when compared with the positive and negative controls, however only the difference between the negative control and the group vaccinated with rDnaK was significant (Fig. 7B). The eosinophil count showed no statistical difference between all groups (Fig. 7C).
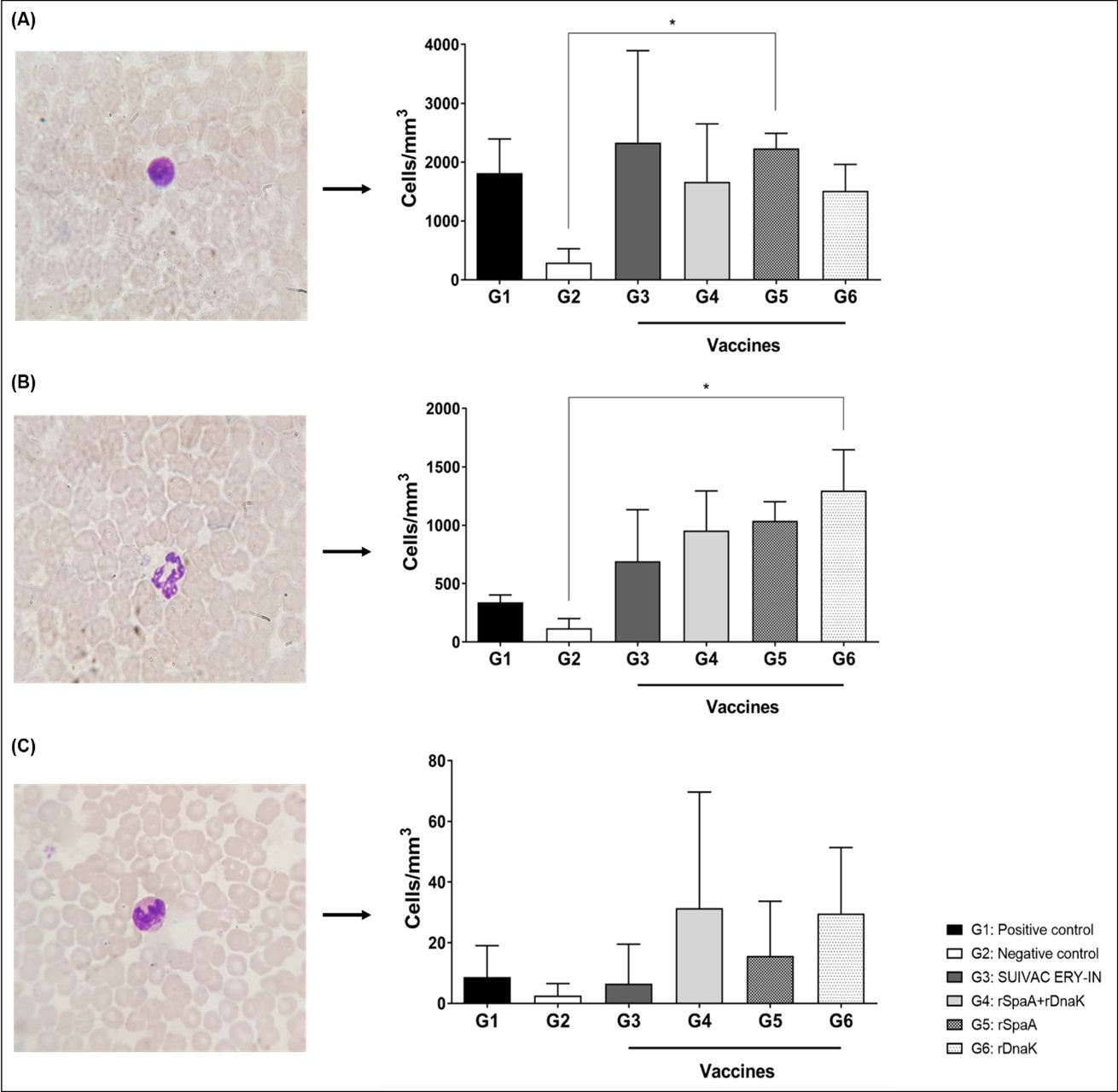
Fig. 7: Morphology and count of leukocytes in the blood of 4 isogenic mice from all 6 groups, 3 days after infection. The slides were made through blood smear and the leukocytes were differentiated according to their morphological varieties and amont in peripheral blood. (A) Morphology and profile of mononuclear cells (B) Morphology and profile of neutrophils. (C) Morphology and profile of eosinophils. *p<0.05 represents the significant difference between the groups.
Colonization of spleen and kidney by E. rhusiopathiae after mice immunization
E. rhusiopathiae biodistribution analysis was performed in the spleens and right kidneys to evaluate the colonization of these organs by the bacterium in the different groups studied. As expected, the group G2 that was challenged with the bacteria without previous vaccination presented a greater number of colonies formed from both analyzed tissues. The group immunized with SUIVAC did not show colony growth, while the rSpaA group showed a minimum growth of 1 to 2 colonies in both organs. Groups vaccinated with rDnaK or rSpaA+rDnaK mixture showed colony growth, but in a smaller number than the observed for the negative control (Fig. 8).
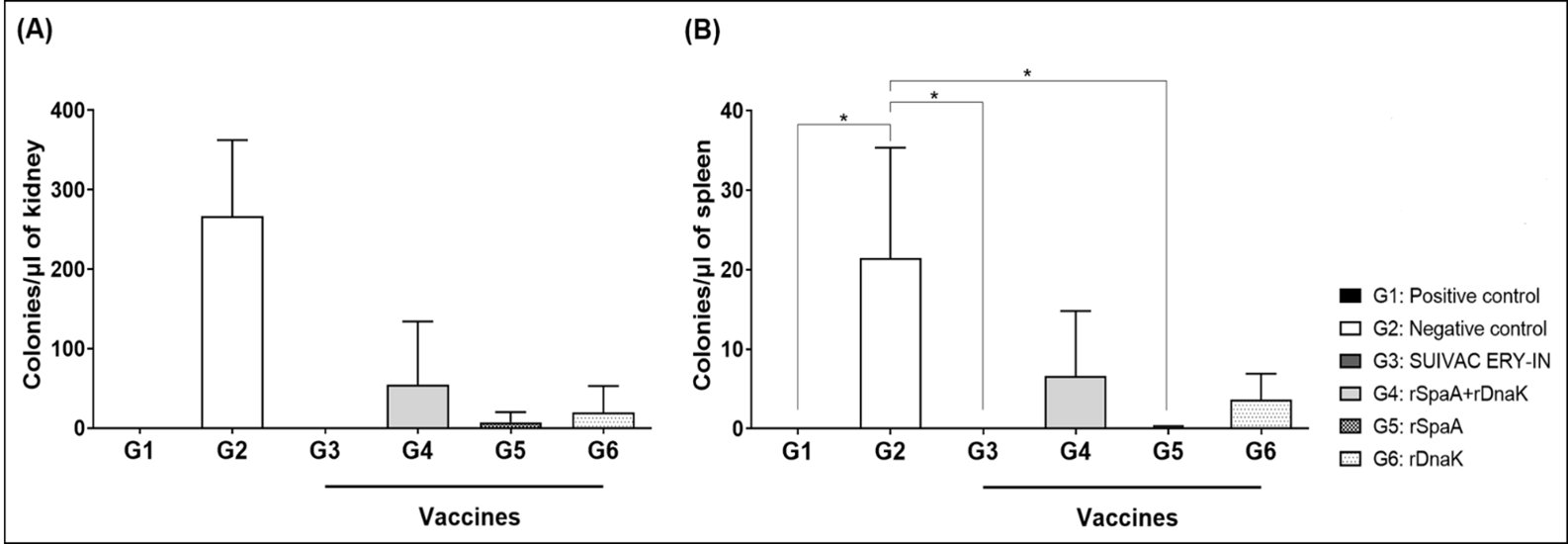
Fig. 8: Biodistribution of kidney and spleen organs from heterogeneous animals, 3 days after infection with 4000 E. rhusiopathiae cells. (A) 100 µL of macerated kidney sample was plated and diluted to 102 in 20 mL of solid LB medium. (B) 100µL of spleen sample macerated and diluted to 101 in 20 mL of solid LB medium. Organs were collected from 3 animals of all six groups tested. The data was normalized and *p<0.05 represents the significant difference between the groups.
Protective effect of recombinant proteins against erysipelas
For the assay of protection against erysipelas, during the period of 14 days the appearance of symptoms such as fever, weakness, closed eyes and apathy, of each group were monitored. The non-vaccinated and challenged mice (G2 group) presented the initial symptoms of the disease at the third day after the challenge, and all animals in the group died on the fifth day. Whereas mice from the G2 group died within 5 days, those from the G4 group, immunized with the two recombinant proteins, presented a delay in the disease symptoms for 60% of the mice, or even a complete protection for 20% of them which did not show any symptoms. Group G5 showed a protective effect for 80% of mice (only 20% died). Group G6 successfully immunized 60% of mice (40% died), however, the onset of symptoms in 1 animal was delayed and was only observed on the eighth day after the challenge (Fig. 9). All mice that died showed weakness, stooped posture, reduced mobility, compromised coat (fever) and partially closed eyes 1 to 2 days before death.
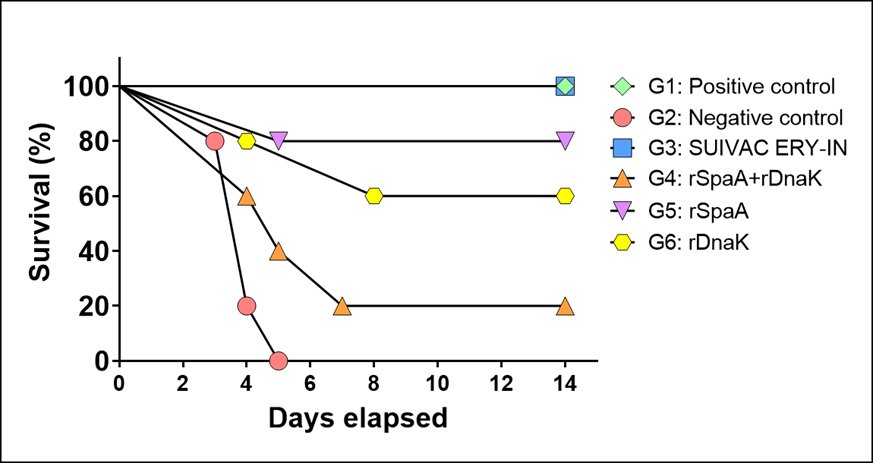
Fig. 9: Survival curve of mice. The survival time of mice was monitored for 14 days from the challenge with 100 µL of 4000 E. rhusiopathiae NCTC11002 cells (day 0), the cells were inoculated subcutaneous and the Positive control received only 1X PBS. In the legend, the Positive control was neither immunized nor challenged; the Negative control was only challenged and the other groups represent the immunogenic agent used in mice that were later also challenged with the bacteria.
Discussion
The search for immunogenic targets that can contribute to vaccine development has led to innovative studies and tools to identify promising proteins. In this study, SpaA and DnaK proteins from E. rhusiopathiae were expressed in E. coli for evaluation of their potential as subunit vaccine for the control of swine erysipelas, both separated or together for the immunization.
The DnaK protein of E. rhusiopathiae is a Hsp70 molecular chaperone with ATPase activity which we previously identified as an immunogenic protein through an immunoproteomic analysis performed to detect E. rhusiopathiae extracellular antigenic proteins that could be potential candidates for subunit vaccine production (unpublished results). The surface protective antigen SpaA is already known in the literature as a protective antigen against E. rhusiopathiae , in addition to have an important role in the bacterial virulence and to be responsible for inducing a high immune response in the host [32, 34, 35]. In Japan, rSpaA is already used as a subunit vaccine for swine under the trade names SUIMMUGEN®rART2/ER (KM Biologics Co., Ltd.) and Swivac ERA (Kyoritsu Seiyaku Corp.) [36]. For swine erysipelas there is no vaccine that is totally protective.
For swine erysipelas, there are reports of mice immunization using E. rhusiopathiae proteins such as CbpB [37], GAPDH [38] and the surface protein SpaA. Imada et al [33]. and To & Nagai [39] respectively tested 50 µg and 40 µg of SpaA protein for their immunization assays, all using the subcutaneous access route. Borrathybay et al [35]. used 100 µg of SpaA protein (subcutaneously) to analyze the immunization degree of this protein. For our immunization assays, each mouse was immunized intramuscular with 100 µg of total protein. Challenge was performed with 4000 cells of the bacterium, since the average lethal dose (LD50) corresponds to 1000 cells.
Humoral and cell-mediated immunity play an important role in host defense against E. rhusiopathiae infection, but humoral immunity plays a critical role, as antiserum treatment provides disease control [40]. In addition, it has been reported that pigs with a few weeks of age are less susceptible to infection due to ingestion of colostrum, which has a high concentration of maternal IgG [41]. Therefore, in this work one of the first characteristics evaluated was the induction of specific IgG antibodies to rSpaA and rDnaK.
The higher titer of IgG against the recombinant proteins was detected in the mice serum inoculated with the recombinant proteins, in comparison to the controls. This indicates that the purified rSpaA and rDnaK were immunogenic, generating a specific humoral response. These data corroborate the study by Cheun et al [42]. and Zhu et al [38]. , who showed a high production of IgG antibodies against the rSpaA protein in serum from mice immunized with the SpaA protein loaded in a live vector Lactococcus lactis or as purified protein, respectively. Shimoji et al [34]. also reported a high production of anti-rSpaA IgG antibodies in the serum of pigs immunized with two doses of an attenuated strain of E. rhusiopathiae , when compared to the serum of pigs immunized with only one dose, and also in serum of non-immunized pigs.
Other studies have reported the protective activity of DnaK in other bacteria, such as the works by Delpino et al [43]. and Hu et al [44]. who demonstrated protective activity of bivalent vaccines, composed with DnaK of Brucella and Edwardsiella tarda , against the respective diseases caused by these microorganisms.
The increase of specific IgG against the proteins indicates that they were antigenic, whereas a change of IgG subclass, to IgG1 or IgG2a, may indicate that each protein induced the immune response of the animals in different ways. We can relate the relative levels of IgG subclasses to the pattern in the immune response of T lymphocytes, Th1 or Th2, and to the interleukins produced by the challenged mice. The switch to IgG1 is induced by the interleukin IL-4, which also inhibits the switch to IgG2a, and is responsible for stimulating a humoral (Th2) response. The switch to IgG2a and the inhibition of the subclass switch to IgG1 are stimulated by interferon-gamma (IFN-γ), which is also responsible for inducing a cell-mediated (Th1) immune response [45, 46].
Analyzing different subclasses of antibodies, IgG1 and IgG2a, rDnaK induced higher levels of IgG1, which is the most abundant subclass and is normally related to responses to viruses, soluble protein and membrane antigens, while rSpaA stimulated a higher level of IgG2a, responsible for responses against bacterial capsular polysaccharides and, less frequently, against protein antigens [47, 48].
A virulence factor of E. rhusiopathiae is associated with resistance to phagocytosis by neutrophils and macrophages, and this ability is closely related to the presence of a polysaccharide capsule on its cell surface [5, 49, 50]. Therefore, it was analyzed how the tested vaccines modulated the immune system of the vaccined mice. rSpaA provided a greater amount of mononucleated cells (monocytes and lymphocytes) when compared to challenged and non-immunized mice. These monocytes are responsible for the phagocytosis of invading bacteria, and the lymphocytes form antibodies and memory cells necessary for the constitution of adaptive immunity [51]. The protein rDnaK provided an increase in neutrophils in the peripheral blood of immunized animals also when compared to challenged and non-immunized mice, being the neutrophils responsible for the phagocytosis of pathogens as well as monocytes, but they remain circulating in the blood for a short time [51].
The biodistribution of E. rhusiopathiae in the spleen and right kidney of mice confirmed that the rSpaA and rDnaK proteins helped the immune system to prevent the proliferation of the bacteria in these organs. However, rSpaA was more efficient as it did not allow the bacteria proliferate in the kidney and spleen, since no CFU was observed in the plaques from the macerated organs. Also in this experiment, the recombinant proteins were more effective when inoculated separately, as the mice vaccinated with the proteins used together (rSpaA+rDnaK) showed a greater number of colonies that grew and were distributed in analyzed organs.
An analysis was also performed to verify the protective activity of rSpaA, rDnaK and rSpaA+rDnaK proteins. In the groups tested, the rSpaA protein provided greater protection to mice when compared with rDnaK and with the two proteins together (rSpaA+rDnaK), protecting 80% of animals against erysipelas. This indicates that SpaA is a promising protein for the composition of a subunit vaccine, as already reported in the work of To & Nagai [39]; Borrathybay [35] and Zhu [38], in which SpaA promoted 100% protection of mice challenged with E. rhusiopathiae. Furthermore, the two proteins when inoculated together (rSpaA+rDnaK) provided the lowest protective effect analyzed in this work, protecting only 20% of the mice. On the other hand, rDnaK induced a partial protective effect but delayed the onset of disease symptoms and the death of the animals, showing that it protected more when compared with the two proteins used together.
Although humoral immunity is very important for host defense, phagocytosis by mononucleates also plays a key role in the elimination of E. rhusiopathiae [50, 52]. We can also observe this importance in this work, since the protein that stimulated more mononucleates, rSpaA, presented a higher percentage of protected animals and a smaller amount of CFU isolated from the kidney and spleen, than rDnaK. These data corroborate the higher amount of IgG2a induced by rSpaA, which suggests a greater induction of the Th1 response than that induced by rDnaK, activating the cellular response and macrophage stimulation.
It is important to emphasize that the protective characterization of rDnaK was performed for the first time in this study, which delayed the onset of symptoms and fully protected 60% of vaccinated and challenged animals, proving to be a promising new protein for the constitution of a subunit vaccine against E. rhusiopathiae . Thus, this study shows that effective protective immunity is a set of events that occur during the process of antigen presentation, effector cells against antigens and the production of antibodies capable of controlling the infection.
It was demonstrated in this work that the DnaK protein from E. rhusiopathiae has immunogenic activity, inducing the production of IgG antibodies in mice. This protein also promotes partial protection, and delays the symptoms of erysipelas in challenged mice, but it is less efficient than the protection provided by SpaA. Proteins were more effective as a vaccine when used separately rather than together, indicating that one does not enhance the protective effect of the other. The SpaA protein is already mentioned in the literature, but this study showed DnaK as an antigenic protein with potential for the formation of a subunit vaccine against swine erysipelas, as it promoted a protective activity in the animals tested.
Abbreviations
CEUA ((Ethics Committee for Animals Use); CFU (Colony Forming Unit); CONCEA (National Council for Control of Animal Experimentation); CV (Column volume); DTT (DL-Dithiothreitol); ELISA (Enzyme-Linked Immunosorbent Assay); His-Tag (6-histidine tag); IMAC (immobilized metal ion affinity chromatography); IPTG (Isopropyl β-D-1-thiogalactopyranoside); LB (Luria Bertani); LD50 (Average lethal dose); PBS (Phosphate-buffered saline); PMSF (Phenylmethylsulfonyl fluoride); SDS-PAGE (Sodium dodecyl sulfate–polyacrylamide gel electrophoresis); SEM (Standard error of the mean); SPF (Specific pathogen-free); TB (Terrific Broth); TMB (3, 3′,5, 5′ tetramethylbenzidine); UFSCar (Federal University of São Carlos).).
Acknowledgements
We thank Dulce Helena Ferreira de Souza (Chemistry Department – UFSCar) for help and lab suport for DnaK exclusion chromatography, and Vinícius Marquioni Monteiro for their help in this work.
The authors acknowledge the support provided by the Brazilian funding agencies: Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES -www.capes.gov.br/) [Finance Code 001] and Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP -www.fapesp.br/) [grant numbers: Nº 18/13465-5, 17/24832-6, and 2014/03003-3, JP-FAPESP 07/50910-2, and CEPID 13/07296-2]. The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
We too thank the ethics c ommittee, CEUA, which involves the production, maintenance and/or use of animals belonging to the phylum Chordata, subphylum Vertebrata (except human beings), for scientific research purposes or teaching - is in accordance with Law 11.794 of October 8, 2008, Decree 6899 of July 15, 2009, as well as with the rules issued by the National Council for Control of Animal Experimentation (CONCEA), and was approved by the CEUA/UFSCAR nº1247201120 in the meeting of 12/04/2020.
Author Contributions
Conceptualization, Godoy, Novo-Mansur and Anibal; methodology, Godoy, Moraes, Zangirolami and da Silva; validation, Godoy, da Silva, Novo-Mansur and Anibal; formal analysis, Godoy, Novo-Mansur and Anibal; investigation, Godoy, Moraes and Santana; resources, Zangirolami, da Silva, Novo-Mansur and Anibal; data curation, Godoy and de Castro; writing-original draft preparation, Godoy; writing-review and editing, Godoy, de Castro, Santana, da Silva, Novo-Mansur and Anibal; visualization, all authors; supervision, Novo-Mansur and Anibal; funding acquisition, Novo-Mansur and Anibal. All authors have read and agreed to the published version of the manuscript.
Disclosure Statement
The authors have no conflicts of interest to declare.
References
| 1 | FAO: http://www.agricultura.gov.br/animal/especies/suinos, 2017,
|
| 2 | ABCS: Panorama da Produção de Suínos no Brasil e no Mundo; Produção de Suínos: Teoria e Prática. Brasília, DF, 2014
|
| 3 | L WR: Erysipelas; Diseases of Swine. USA: Iowa State University Press, 1999
|
| 4 | Reboli AC, Farrar WE: Erysipelothrix rhusiopathiae: an occupational pathogen. Clin Microbiol Rev 1989;2:354-359.
https://doi.org/10.1128/CMR.2.4.354 |
| 5 | Shimoji Y, Yokomizo Y, Mori Y: Intracellular survival and replication of Erysipelothrix rhusiopathiae within murine macrophages: failure of induction of the oxidative burst of macrophages. Infection and immunity 1996;64:1789-1793.
https://doi.org/10.1128/iai.64.5.1789-1793.1996 |
| 6 | Shimoji Y, Ogawa Y, Osaki M, Kabeya H, Maruyama S, Mikami T, Sekizaki T: Adhesive surface proteins of Erysipelothrix rhusiopathiae bind to polystyrene, fibronectin, and type I and IV collagens. J Bacteriol 2003;185:2739-2748.
https://doi.org/10.1128/JB.185.9.2739-2748.2003 |
| 7 | Krasemann C, Muller HE: [The virulence of erysipelothrix rhusiopathiae strains and their neuraminidase production (author's transl)]. Zentralbl Bakteriol Orig A 1975;231:206-213.
|
| 8 | Oliveira SJ: Swine erysipelas: an important problem to pig production. Acta Scientiae Veterinariae 2009;37:97-104.
|
| 9 | Yamamoto K, Kijima M, Yoshimura H, Takahashi T: Antimicrobial susceptibilities of Erysipelothrix rhusiopathiae isolated from pigs with swine erysipelas in Japan, 1988-1998. J Vet Med B Infect Dis Vet Public Health 2001;48:115-126.
https://doi.org/10.1046/j.1439-0450.2001.00427.x |
| 10 | Takahashi T, Sawada T, Muramatsu M, Tamura Y, Fujisawa T, Benno Y, Mitsuoka T: Serotype, antimicrobial susceptibility, and pathogenicity of Erysipelothrix rhusiopathiae isolates from tonsils of apparently healthy slaughter pigs. J Clin Microbiol 1987;25:536-539.
https://doi.org/10.1128/jcm.25.3.536-539.1987 |
| 11 | Imada Y, Takase A, Kikuma R, Iwamaru Y, Akachi S, Hayakawa Y: Serotyping of 800 strains of Erysipelothrix isolated from pigs affected with erysipelas and discrimination of attenuated live vaccine strain by genotyping. J Clin Microbiol 2004;42:2121-2126.
https://doi.org/10.1128/JCM.42.5.2121-2126.2004 |
| 12 | Gerber PF, MacLeod A, Opriessnig T: Erysipelothrix rhusiopathiae serotype 15 associated with recurring pig erysipelas outbreaks. Vet Rec 2018;182:635.
https://doi.org/10.1136/vr.104421 |
| 13 | Opriessnig T, Hoffman LJ, Harris DL, Gaul SB, Halbur PG: Erysipelothrix rhusiopathiae: genetic characterization of midwest US isolates and live commercial vaccines using pulsed-field gel electrophoresis. J Vet Diagn Invest 2004;16:101-107.
https://doi.org/10.1177/104063870401600202 |
| 14 | Bender JS, Shen HG, Irwin CK, Schwartz KJ, Opriessnig T: Characterization of Erysipelothrix species isolates from clinically affected pigs, environmental samples, and vaccine strains from six recent swine erysipelas outbreaks in the United States. Clinical and vaccine immunology : CVI 2010;17:1605-1611.
https://doi.org/10.1128/CVI.00206-10 |
| 15 | Zou Y, Zhu X, Muhammad HM, Jiang P, Li Y: Characterization of Erysipelothrix rhusiopathiae strains isolated from acute swine erysipelas outbreaks in Eastern China. J Vet Med Sci 2015;77:653-660.
https://doi.org/10.1292/jvms.14-0589 |
| 16 | To H, Sato H, Tazumi A, Tsutsumi N, Nagai S, Iwata A, Nagano T: Characterization of Erysipelothrix rhusiopathiae strains isolated from recent swine erysipelas outbreaks in Japan. J Vet Med Sci 2012;74:949-953.
https://doi.org/10.1292/jvms.11-0533 |
| 17 | Uchiyama M, Yamamoto K, Ochiai M, Yamamoto T, Hirano F, Imamura S, Nagai H, Ohishi K, Horiuchi N, Kijima M: Prevalence of Met-203 type spaA variant in Erysipelothrix rhusiopathiae isolates and the efficacy of swine erysipelas vaccines in Japan. Biologicals 2014;42:109-113.
https://doi.org/10.1016/j.biologicals.2013.12.002 |
| 18 | Morimoto M, Kato A, Kojima H, Akaike Y, Nogami K, Sasakawa C, Nagai S, To H: Serovars and SpaA Types of Erysipelothrix rhusiopathiae Isolated from Pigs in Japan from 2012 to 2019. Curr Microbiol 2021;78:55-66.
https://doi.org/10.1007/s00284-020-02254-2 |
| 19 | Neumann EJ, Grinberg A, Bonistalli KN, Mack HJ, Lehrbach PR, Gibson N: Safety of a live attenuated Erysipelothrix rhusiopathiae vaccine for swine. Vet Microbiol 2009;135:297-303.
https://doi.org/10.1016/j.vetmic.2008.09.059 |
| 20 | McNeil M, Gerber PF, Thomson J, Williamson S, Opriessnig T: Serotypes and Spa types of Erysipelothrix rhusiopathiae isolates from British pigs (1987 to 2015). Vet J 2017;225:13-15.
https://doi.org/10.1016/j.tvjl.2017.04.012 |
| 21 | Liljeqvist S, Stahl S: Production of recombinant subunit vaccines: protein immunogens, live delivery systems and nucleic acid vaccines. Journal of biotechnology 1999;73:1-33.
https://doi.org/10.1016/S0168-1656(99)00107-8 |
| 22 | Ingebritson AL, Roth JA, Hauer PJ: Erysipelothrix rhusiopathiae: association of Spa-type with serotype and role in protective immunity. Vaccine 2010;28:2490-2496.
https://doi.org/10.1016/j.vaccine.2010.01.041 |
| 23 | Makino SI, Yamamoto K, Asakura H, Shirahata T: Surface antigen, SpaA, of erysipelothrix rhusiopathiae binds to Gram-positive bacterial cell surfaces. FEMS Microbiol Lett 2000;186:313-317.
https://doi.org/10.1111/j.1574-6968.2000.tb09123.x |
| 24 | Partridge J, King J, Krska J, Rockabrand D, Blum P: Cloning, heterologous expression, and characterization of the Erysipelothrix rhusiopathiae DnaK protein. Infection and immunity 1993;61:411-417.
https://doi.org/10.1128/iai.61.2.411-417.1993 |
| 25 | Suzue K, Young RA: Heat shock proteins as immunological carriers and vaccines. EXS 1996;77:451-465.
https://doi.org/10.1007/978-3-0348-9088-5_30 |
| 26 | da Silva AJ, da Costa Iemma MR, Luperni Horta AC, Sargo CR, de Lima Camargo Giordano R, de Campos Giordano R, Zangirolami TC, Marques Novo MT: Cloning, auto-induction expression, and purification of rSpaA swine erysipelas antigen. Current microbiology 2012;65:369-374.
https://doi.org/10.1007/s00284-012-0171-y |
| 27 | Hochuli E: Large-scale chromatography of recombinant proteins. J Chromatogr 1988;444:293-302.
https://doi.org/10.1016/S0021-9673(01)94032-4 |
| 28 | Laemmli UK: Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970;227:680-685.
https://doi.org/10.1038/227680a0 |
| 29 | Bradford MM: A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 1976;72:248-254.
https://doi.org/10.1016/0003-2697(76)90527-3 |
| 30 | Chernecky CC, Berger BJ: Laboratory Tests and Diagnostic Procedures. Elsevier Saunders, 2013.
|
| 31 | Kaplan EL, Meier, P.: Nonparametric Estimation from Incomplete Observations. Journal of the American Statistical Association 1958;53:457-481.
https://doi.org/10.1080/01621459.1958.10501452 |
| 32 | Makino S, Yamamoto K, Murakami S, Shirahata T, Uemura K, Sawada T, Wakamoto H, Morita H: Properties of repeat domain found in a novel protective antigen, SpaA, of Erysipelothrix rhusiopathiae. Microbial pathogenesis 1998;25:101-109.
https://doi.org/10.1006/mpat.1998.0216 |
| 33 | Imada Y, Goji N, Ishikawa H, Kishima M, Sekizaki T: Truncated surface protective antigen (SpaA) of Erysipelothrix rhusiopathiae serotype 1a elicits protection against challenge with serotypes 1a and 2b in pigs. Infection and immunity 1999;67:4376-4382.
https://doi.org/10.1128/IAI.67.9.4376-4382.1999 |
| 34 | Shimoji Y, Oishi E, Kitajima T, Muneta Y, Shimizu S, Mori Y: Erysipelothrix rhusiopathiae YS-1 as a live vaccine vehicle for heterologous protein expression and intranasal immunization of pigs. Infection and immunity 2002;70:226-232.
https://doi.org/10.1128/IAI.70.1.226-232.2002 |
| 35 | Borrathybay E, Gong FJ, Zhang L, Nazierbieke W: Role of surface protective antigen A in the pathogenesis of Erysipelothrix rhusiopathiae strain C43065. J Microbiol Biotechnol 2015;25:206-216.
https://doi.org/10.4014/jmb.1407.07058 |
| 36 | Opriessnig T, Forde T, Shimoji Y: Erysipelothrix Spp.: Past, Present, and Future Directions in Vaccine Research. Front Vet Sci 2020;7:174.
https://doi.org/10.3389/fvets.2020.00174 |
| 37 | Shi F, Ogawa Y, Sano A, Harada T, Hirota J, Eguchi M, Oishi E, Shimoji Y: Characterization and identification of a novel candidate vaccine protein through systematic analysis of extracellular proteins of Erysipelothrix rhusiopathiae. Infection and immunity 2013;81:4333-4340.
https://doi.org/10.1128/IAI.00549-13 |
| 38 | Zhu W, Wu C, Kang C, Cai C, Wang Y, Li J, Zhang Q, Sun X, Jin M: Evaluation of the protective efficacy of four newly identified surface proteins of Erysipelothrix rhusiopathiae. Vaccine 2018;36:8079-8083.
https://doi.org/10.1016/j.vaccine.2018.10.071 |
| 39 | To H, Nagai S: Genetic and antigenic diversity of the surface protective antigen proteins of Erysipelothrix rhusiopathiae. Clinical and vaccine immunology : CVI 2007;14:813-820.
https://doi.org/10.1128/CVI.00099-07 |
| 40 | Yokomizo Y, Isayama Y: Antibody activities of IgM and IgG fractions from rabbit anti-Erysipelothrix rhusiopathiae sera. Res Vet Sci 1972;13:294-296.
https://doi.org/10.1016/S0034-5288(18)34048-7 |
| 41 | Imada Y, Mori Y, Daizoh M, Kudoh K, Sakano T: Enzyme-linked immunosorbent assay employing a recombinant antigen for detection of protective antibody against swine erysipelas. J Clin Microbiol 2003;41:5015-5021.
https://doi.org/10.1128/JCM.41.11.5015-5021.2003 |
| 42 | Cheun HI, Kawamoto K, Hiramatsu M, Tamaoki H, Shirahata T, Igimi S, Makino SI: Protective immunity of SpaA-antigen producing Lactococcus lactis against Erysipelothrix rhusiopathiae infection. Journal of applied microbiology 2004;96:1347-1353.
https://doi.org/10.1111/j.1365-2672.2004.02283.x |
| 43 | Delpino MV, Estein SM, Fossati CA, Baldi PC, Cassataro J: Vaccination with Brucella recombinant DnaK and SurA proteins induces protection against Brucella abortus infection in BALB/c mice. Vaccine 2007;25:6721-6729.
https://doi.org/10.1016/j.vaccine.2007.07.002 |
| 44 | Hu YH, Dang W, Deng T, Sun L: Edwardsiella tarda DnaK: expression, activity, and the basis for the construction of a bivalent live vaccine against E. tarda and Streptococcus iniae. Fish Shellfish Immunol 2012;32:616-620.
https://doi.org/10.1016/j.fsi.2012.01.011 |
| 45 | Snapper CM, Paul WE: Interferon-gamma and B cell stimulatory factor-1 reciprocally regulate Ig isotype production. Science 1987;236:944-947.
https://doi.org/10.1126/science.3107127 |
| 46 | Stevens TL, Bossie A, Sanders VM, Fernandez-Botran R, Coffman RL, Mosmann TR, Vitetta ES: Regulation of antibody isotype secretion by subsets of antigen-specific helper T cells. Nature 1988;334:255-258.
https://doi.org/10.1038/334255a0 |
| 47 | Ferrante A, Beard LJ, Feldman RG: IgG subclass distribution of antibodies to bacterial and viral antigens. Pediatr Infect Dis J 1990;9:S16-24.
https://doi.org/10.1097/00006454-199008001-00004 |
| 48 | Vidarsson G, Dekkers G, Rispens T: IgG subclasses and allotypes: from structure to effector functions. Front Immunol 2014;5:520.
https://doi.org/10.3389/fimmu.2014.00520 |
| 49 | Shimoji Y, Yokomizo Y, Sekizaki T, Mori Y, Kubo M: Presence of a capsule in Erysipelothrix rhusiopathiae and its relationship to virulence for mice. Infection and immunity 1994;62:2806-2810.
https://doi.org/10.1128/iai.62.7.2806-2810.1994 |
| 50 | Timoney J: The effect of decomplementation on Erysipelothrix rhusiopathiae infection in the mouse. Immunology 1970;19:561-567.
|
| 51 | Abbas AK, Lichtman AH, Pillai S: Cellular and Molecular Immunology Elsevier, 2017.
|
| 52 | Timoney J: The inactivation of Erysipelothrix rhusiopathiae in pig buffy-coat leucocytes. Res Vet Sci 1970;11:189-190.
https://doi.org/10.1016/S0034-5288(18)34359-5 |